Radiologia Brasileira - Publicação Científica Oficial do Colégio Brasileiro de Radiologia
AMB - Associação Médica Brasileira CNA - Comissão Nacional de Acreditação
 Vol. 41 nº 1 - Jan. / Fev. of 2008
Vol. 41 nº 1 - Jan. / Fev. of 2008
|
ARTIGO ORIGINAL
|
|
|
|
|
Autho(rs): Liliane Lins, Laís Gomes, Lis Gomes, Marcele Trindade, Leonardo Dias, Ricardo Bragança, Rodrigo Pimentel |
|
|
Descritores: Irradiação, Malformações, Hemorragia, Sistema nervoso central |
|
|
Resumo:
INTRODUÇÃO Neuroblastos, provavelmente o mais abundante tipo de célula presente nos fetos de mamíferos, são muito radiossensíveis e representam um estágio intermediário entre as células neuroepiteliais e neurônios. Em camundongos, os neuroblastos aparecem sete dias após a concepção, enquanto na espécie humana são formados 18 dias após a fecundação. Tanto na espécie humana quanto em camundongos essas células estão presentes desde a gestação até o período neonatal, formando conexões com tecidos em desenvolvimento e órgãos(1). Sendo assim, o uso de radiação no período embrionário, quando os neuroblastos encontram-se mais concentrados, freqüentemente resulta em anomalias congênitas do sistema nervoso central e órgãos associados, tais como microftalmia, anoftalmia, microcefalia e anencefalia. Estudos demonstraram que uma dose de algumas centenas de Gy de raios-X pode induzir significante número de hemorragias no encéfalo, sendo a quantidade de lesões exponencialmente proporcional à elevação da dose(2). Entretanto, ainda há a necessidade de análise dos efeitos de baixas doses de radiação-X no desenvolvimento do sistema nervoso de fetos de ratos. O efeito da radiação na fase de pré-implantação pode ser mais bem descrito como "tudo ou nada", dicotomicamente representado pela morte precoce do embrião ou pelo seu desenvolvimento normal. Considera-se que o dano cromossomal causado pela irradiação seja o mecanismo mais importante responsável pela morte do embrião, pois promove a degeneração de células primitivas. O principal efeito da exposição à radiação na organogênese é o desenvolvimento de malformações(3). As anomalias induzidas dependem dos órgãos sofrendo organogênese no período da irradiação, do estágio da diferenciação e da dose aplicada(4). Déficits graves e persistentes no cérebro adulto, arquitetura cortical desorganizada, redução do tamanho cortical e do peso cerebral, microcefalia e disfunção motora foram observadas no estudo de Miki et al.(5), que aplicaram irradiações sucessivas, do 13º ao 18º dia gestacional, de raios-X em doses de 1,0 Gy a 2,0 Gy. Takai et al.(6) expuseram ratos adultos a uma dose de radiação-X de 1,5 Gy no hipocampo, resultando em disfunção cognitiva, relacionada à ectopia de células. Ainda em relação às alterações hipocampais, Schmitz et al.(7), trabalhando com radiação gama, relataram diminuição de células de Purkinje e aumento volumétrico como resultado da aplicação de uma dose de 3,0 Gy, do 13º ao 16º dias gestacionais. Devido à necessidade de mais estudos histomorfológicos que caracterizem os efeitos dos raios-X no desenvolvimento embrionário, o objetivo do presente estudo é analisar os efeitos de baixas doses de radiação-X no desenvolvimento do sistema nervoso de fetos de ratos.
MATERIAIS E MÉTODOS O presente estudo foi constituído de 15 Rattus norvegicus albinos, Wistar, sendo dez fêmeas e cinco machos com, aproximadamente, oito semanas de idade e peso médio de 275 gramas. Machos e fêmeas foram mantidos em um ciclo de 12 horas claro/escuro e tiveram acesso irrestrito a água e comida. As fêmeas foram fertilizadas de acordo com uma adaptação do método de Chahoud e Kwasigroch(8). Para a detecção do tampão vaginal, foi utilizado um otoscópio, estabelecendo-se o dia da concepção(9). Cinco fêmeas no oitavo dia gestacional, com 37 embriões, foram mantidas em padrões normais de sobrevivência, compondo o grupo controle. Outras cinco fêmeas, também no oitavo dia gestacional, com 39 embriões, foram expostas a uma única dose de raios-X, de 0,3 Gy, por 30 segundos, na região abdominal. Foi usado, para a irradiação, um aparelho odontológico Gnatus, de 70 kV e 10 mA. Todos os ratos foram confinados em gaiolas separadas e anestesiados com éter para os procedimentos de irradiação e perfusão. No 17º dia de gestação, ambos os grupos foram submetidos a histerectomia e sacrificados com injeção de éter na cavidade abdominal, sob anestesia de ketamina 0,2 ml/100 g. Após a histerectomia, os úteros dos animais de ambos os grupos foram fixados em formol a 10%, desidratados e embebidos em parafina. Várias secções de 4 µm foram feitas com um micrótomo. Secções selecionadas foram coradas com hematoxilina-eosina e examinadas para análise comparativa do encéfalo dos fetos de ambos os grupos. A análise macroscópica foi baseada no fechamento do neuróporo rostral e a análise microscópica consistiu em avaliação morfológica do encéfalo utilizando magnificação de dez vezes. As áreas analisadas foram hipocampo e cerebelo, além da ectopia neuronal, perda da arquitetura do tecido e hemorragia, sendo esta classificada em leve, moderada e severa, de acordo com a intensidade da congestão dos vasos e o volume de hemácias extravasado. Para a análise estatística utilizou-se o programa Epi-Info 2004 (CDC, versão 3.3), de acordo com o teste do qui-quadrado para as variáveis hemorragia e alteração da arquitetura do tecido nervoso, comparando os grupos controle e experimental. Os resultados foram considerados estatisticamente significantes quando p < 0,05.
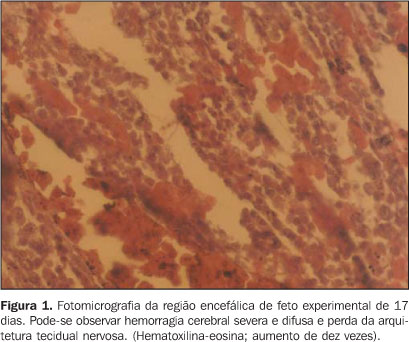
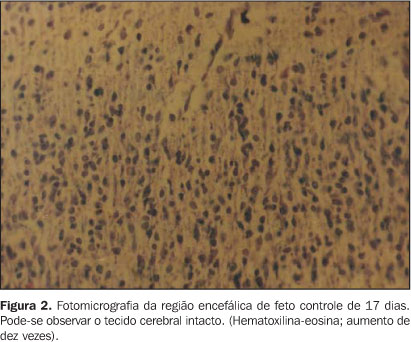
RESULTADOS Foram analisados 37 e 39 embriões, respectivamente, nos grupos controle e experimental, não ocorrendo perda de seguimento. Exames clínicos revelaram ausência de diferenças macroscópicas entre os grupos controle e experimental, uma vez que 100% dos animais obtiveram fechamento normal do neuróporo rostral e nenhum deles (0%) apresentou sinais como anoftalmia, microcefalia e anencefalia. Análises microscópicas revelaram hemorragia do córtex cerebral do grupo experimental, variando entre moderada a severa. Vinte e sete por cento dos animais do grupo experimental apresentaram hemorragia cerebral moderada e 77% apresentaram hemorragia severa (p < 0,05; Figura 1). Os animais com hemorragia severa também apresentaram perda da arquitetura do tecido nervoso (p = 0,05). Nenhum animal do grupo controle apresentou hemorragia cerebral ou perda de arquitetura do tecido nervoso (Figura 2).
DISCUSSÃO Todas as gestações evoluíram normalmente até o período da análise, não tendo sido observada nenhuma alteração comportamental no grupo experimental, quando comparado ao grupo controle. A detecção do plug vaginal foi realizada de forma efetiva por um otoscópio, o que permitiu a determinação do dia exato da concepção e o aumento da predictabilidade da gravidez, como demonstrado por Voipio e Nevalainen(9). Esse método não causa nenhum estresse ou dor aos animais, é rápido e de fácil realização. Dessa forma, a introdução do otoscópio não apresenta efeitos morfológicos no desenvolvimento da gravidez; por conseguinte, os animais não sofreram interferência externa durante o estudo. Uma vez que as células do tecido nervoso são extremamente sensíveis à radiação, lesões do tecido nervoso e hemorragias foram os principais achados deste estudo. Entretanto, outros efeitos como disfunções motoras e cognitivas, atrofia das células de Purkinje, microcefalia e ectopia hipocampal foram detectadas por outros autores(5,6), contudo, não foram investigadas no presente estudo. As hemorragias detectadas no cérebro de ratos expostos à radiação são evidências consistentes com os achados de Yang e Tobias(2), os quais demonstraram que uma baixa dose de radiação-X pode causar hemorragias cerebrais e, conseqüentemente, lesão tecidual nervosa. Nenhum dos animais apresentou anormalidades como anoftalmia, microcefalia e anencefalia, provavelmente devido à baixa dose aplicada, segundo os achados de Friedberg et al.(4). Apesar do grupo experimental ter apresentado altos níveis de hemorragia cerebral, não houve diferenças morfológicas entre os grupos experimental e controle. Esses achados são consistentes com Wang et al.(10), que encontraram poucas alterações induzidas pela radiação. A descrição do efeito da irradiação no período de pré-implantação como "tudo ou nada"(3) foi confirmada no presente estudo, uma vez que os embriões se desenvolveram normalmente até o 17º dia gestacional, não havendo perda de seguimento. Vos(3) ainda afirma, em seu estudo, que o principal efeito da exposição aos raios-X na fase da organogênese é o desenvolvimento de malformações. Apesar de no presente estudo o grupo experimental ter sido irradiado no período da organogênese, nenhuma malformação foi evidenciada. Contudo, o tecido nervoso com perda da arquitetura e hemorragia provavelmente acarretaria distúrbios de desenvolvimento posteriores. De acordo com Schmitz et al.(7), irradiação pré-natal resulta não somente em perda neuronal, mas também em decréscimo massivo de volume nas regiões cerebrais investigadas. Isso não foi observado neste estudo, provavelmente devido à baixa dose aplicada. Sabe-se que as células progenitoras podem ser lesadas pela radiação e não se dividem durante a exposição, mas talvez uma única dose de irradiação não seja suficiente para causar danos nos estágios de desenvolvimento celular. Esses autores aplicaram graus diferentes de irradiação, o que pode ter sido a causa de interferência nos fatores de crescimento. Em conclusão, os achados do presente trabalho suportam a assertiva de que radiação-X, na forma e condições aqui aplicadas, pode determinar a ocorrência de hemorragias cerebrais e lesões teciduais nervosas em fetos de ratos. Agradecimentos Este trabalho foi em parte apoiado pela Fundação Bahiana para Desenvolvimento das Ciências.
REFERÊNCIAS 1. Rugh R. X-ray-induced teratogenesis in the mouse and its possible significance to man. Radiology. 1971;99:433–43. [ ] 2. Yang T, Tobias C. Effects of heavy ion irradiation on the brain vascular system and embryonic development. Adv Space Res. 1984;4:239–45. [ ] 3. Vos O. Effects and consequences of prenatal irradiation. Boll Soc Ital Biol Sper. 1989;65:481–500. [ ] 4. Friedberg W, Faulkner DN, Neas BR, et al. Dose-incidence relationships for exencephalia, anophthalmia and prenatal mortality in mouse embryos irradiated with fission neutrons or 250 kVX-rays. Int J Radiat Biol Relat Stud Phys Chem Med. 1987;52:223–36. [ ] 5. Miki T, Fukui Y, Takeuchi Y, et al. A quantitative study of the effects of prenatal X-irradiation on the development of cerebral cortex in rats. Neurosci Res. 1995;23:241–7. [ ] 6. Takai N, Sun XZ, Ando K, et al. Ectopic neurons in the hippocampus may be a cause of learning disability after prenatal exposure to X-rays in rats. J Radiat Res. 2004;45:563–9. [ ] 7. Schmitz C, Born M, Dolezel P, et al. Prenatal protracted irradiation at very low dose rate induces severe neuronal loss in rat hippocampus and cerebellum. Neuroscience. 2005;130:935–48. [ ] 8. Chahoud I, Kwasigroch TE. Controlled breeding of laboratory animals In: Neubert D, Merker HJ, Kwasigroch TE, editors. Methods in prenatal toxicology. Stuttgart: Georg Thieme; 1977. p.78–91. [ ] 9. Voipio HM, Nevalainen T. Improved method for vaginal plug detection in rats. Scand J Lab Anim Sci. 1998;25:5–9. [ ] 10. Wang H, Chen D, Gao C, et al. Effects of low level prenatal 60Co gamma-irradiation on postnatal growth and behavior in mice. Teratology. 1993; 48:451–7. [ ]
Recebido para publicação em 15/5/2006. Aceito, após revisão, em 16/8/2007.
* Trabalho realizado na Escola Bahiana de Medicina e Saúde Pública, Salvador, BA, Brasil. |
|

Av. Paulista, 37 - 7° andar - Conj. 71 - CEP 01311-902 - São Paulo - SP - Brazil - Phone: (11) 3372-4544 - Fax: (11) 3372-4554