Radiologia Brasileira - Publicação Científica Oficial do Colégio Brasileiro de Radiologia
AMB - Associação Médica Brasileira CNA - Comissão Nacional de Acreditação
 Vol. 38 nº 6 - Nov. / Dez. of 2005
Vol. 38 nº 6 - Nov. / Dez. of 2005
|
ARTIGO DE REVISÃO
|
|
|
|
|
Autho(rs): Heloisa de Andrade Carvalho |
|
|
Descritores: Radiobiologia, Mutação, Tradescantia, Radiação ionizante |
|
|
Resumo:
INTRODUÇÃO Desde a sua descoberta, em 1896, as radiações ionizantes têm tido utilização crescente, tanto em medicina quanto em outras áreas. Cada vez mais a energia atômica vem sendo utilizada, aumentando a radiação ambiente, podendo levar a um crescimento do risco de mutações em gerações futuras, induzidas pelas radiações. Assim, sistemas de correlação entre doses baixas de radiação e a freqüência de mutações têm sido desenvolvidos de forma a se poder estimar esse risco.
PLANTAS SUPERIORES E BIOMONITORIZAÇÃO Na prática, idealmente, deveriam ser utilizados mamíferos para se estudar os efeitos das radiações e os resultados poderiam ser extrapolados para o ser humano. Entretanto, tais estudos são de difícil realização e requerem uma amostra muito grande, de maneira a se obter resultados consistentes, estatisticamente significantes, de um aumento das mutações em relação à freqüência natural de sua ocorrência. O interesse na utilização de plantas superiores para a detecção de agentes ambientais com potencial mutagênico tem sido crescente. Experimentos com diversos vegetais são descritos, desde 1913(1), quando se utilizou a Vicia faba (cebola) para experimentos com radiação. A Tradescantia é uma planta que apresenta fácil adaptação em qualquer ambiente e pode se desenvolver durante todo o ano, tanto ao ar livre, nas regiões subtropicais, quanto em estufas, em qualquer parte do mundo. O tamanho, relativamente pequeno, e o código genético composto por seis pares de cromossomos relativamente grandes tornaram essa planta um instrumento favorável para estudos citogenéticos.
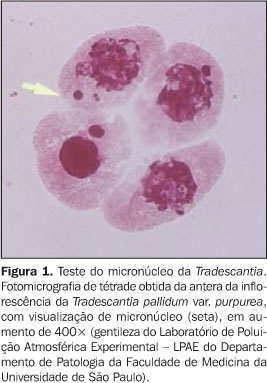
TRADESCANTIA A influência de agentes químicos e físicos (em especial a radiação) sobre a freqüência de mutações tem sido amplamente estudada mediante análise de alterações observadas na Tradescantia. O seu sistema de estame, composto por inúmeros filamentos, é heterozigoto para a cor da flor, sendo azul o dominante e rosa o recessivo. Cada inflorescência formada na extremidade do caule é composta por mais de 50 botões de flores num estágio seqüencial de desenvolvimento, que desabrocham (um ou dois) diariamente. Cada flor é composta por seis estames (três antepétalas e três ante-sépalas) que sustentam de 50 a 120 filamentos. Cada filamento é uma cadeia única de cerca de 30 a 35 células, que são o produto das divisões celulares das células das extremidades, que são meristemáticas, até que o filamento esteja completamente desenvolvido(2,3). A avaliação das alterações genéticas da Tradescantia pode ser feita pela detecção de mutações somáticas induzidas por mutágenos presentes no ar, no solo e na água ou detecção de aberrações cromossômicas induzidas por agentes clastógenos (que promovem a quebra dos cromossomos) presentes no ar, no solo e na água. Para tal, foram desenvolvidos o ensaio do filamento do estame, que pode ser utilizado na avaliação das mutações somáticas, e o teste do tubo de pólen e o ensaio do micronúcleo, desenvolvidos para a detecção de aberrações cromossômicas durante a mitose e meiose, respectivamente. Utilizando clones heterozigotos para a cor da flor, pode-se avaliar a ocorrência de mutação ou perda do gene dominante, facilmente, através da avaliação da cor dos filamentos do estame. A possibilidade de se avaliar uma amostra significante (ao redor de 300 a 700 filamentos ou até 7.500 a 18.000 células dos filamentos em uma única flor), pela simples avaliação de alteração da cor azul para rosa (identificando o índice de células mutantes), favorece esse tipo de ensaio para o estudo de efeitos genéticos de mutágenos em baixas doses de exposição(4). Esse tipo de avaliação ficou conhecido como ensaio do filamento do estame da Tradescantia e foi desenvolvido por Sparrow, do Brookhaven National Laboratory, nos EUA(4,5). Foi explorado como um teste de mutação somática em radiobiologia, mutagênese química e monitoração do ar ambiente(4,6–8). Além disso, estudos extensos sobre os efeitos diretos e indiretos das radiações nos cromossomos mitóticos dos microesoporos da Tradescantia levaram ao estabelecimento de inúmeros princípios na indução de quebras e aberrações cromossômicas induzidas pelas radiações, além da interpretação do cromossomo aberrante e da configuração da cromátide(9–11). Os cromossomos da extremidade da raiz da Tradescantia foram utilizados para o estudo de mutágenos químicos e lesões cromossômicas induzidas por radiação. E os cromossomos das células mitóticas do tubo polínico da Tradescantia foram utilizados no estudo de alterações induzidas pela luz ultravioleta e radiação ionizante até o final de 1940(12–16). A partir de então, foi possível o cultivo de grãos de pólen maduros num meio simples de ágar-lactose que, quando submetidos a um mutágeno químico ou radiação, poderiam apresentar aberrações na cromátide(12–15,17–19). Foi instituído, assim, o teste do tubo de pólen da Tradescantia para a identificação de aberrações cromossômicas durante a mitose. Apesar dos cromossomos das células-mãe germinativas em meiose apresentarem maior sensibilidade à radiação do que os cromossomos em mitose(9,10,20), raramente eram utilizados para o estudo das aberrações induzidas pela radiação. Essas aberrações eram de difícil análise e contagem para se obter dados quantitativos consistentes. Foi desenvolvido, então, o bioensaio do micronúcleo da Tradescantia ("Tradescantia micronucleus bioassay" – Trad-MCN)(21,22), utilizado pela primeira vez por Ma et al. em 1978(23), para a avaliação dos efeitos do 1,2-dibrometano sobre os cromossomos das células em meiose. A base para o desenvolvimento desse ensaio foi o fato de que o maior problema na avaliação quantitativa das aberrações cromossômicas estava na perda dos cromossomos em metáfase I e sua imagem borrada nas preparações das células em meiose. Entretanto, se o agente é aplicado no início da prófase I e o cromossomo prossegue por um período de recuperação, os fragmentos acêntricos dos cromossomos se transformam em micronúcleos, na fase de tétrade da meiose, facilmente identificados à microscopia de luz (Figura 1).
TRADESCANTIA E RADIAÇÕES IONIZANTES Ma, em 1979(24), publicou uma curva dose-resposta para raios-X obtida por ensaio do micronúcleo. Observa-se que a resposta é linear, com aumento do índice de micronúcleos quando se aumenta a dose de radiação. Essa curva, no entanto, é muito pouco precisa quando se leva em conta doses baixas de radiação (< 5 cGy) que estão presentes nos locais de risco. O mesmo tipo de curva já havia sido obtida por Sparrow et al. em 1972(25), inclusive com doses mínimas de até 0,25 cGy, para nêutrons e raios-X, utilizando, entretanto, o método de avaliação do filamento do estame da Tradescantia. Foi observada relação linear entre o índice de mutações e a dose de radiação abaixo de 5 cGy. Com doses entre 10 e 100 cGy, foi observado um componente quadrático na curva de resposta aos raios-X, compatível com um provável reparo de lesões nesses níveis mais elevados de doses. Pode-se concluir que, provavelmente, os efeitos da radiação em doses baixas são devidos essencialmente aos eventos de choque único. Uma série de clones heterozigotos para a cor da flor da Tradescantia (azul/rosa) está disponível para testes de mutagenicidade: BNL 02, KU 27, BNL 4430, KU 9, KU 7, KU 20 e BNL 2031. São classificados de acordo com sua taxa de ocorrência de mutações espontâneas(3). Dependendo dessa característica e sensibilidade aos diversos agentes mutagênicos, são mais adequados para um ou outro tipo de experimento, quer seja em laboratório ou em estudo de campo. Os clones BNL 4430, BNL 02, KU 9 e KU20 são particularmente sensíveis à radiação em graus variados. O clone 4430 da Tradescantia é um diplóide (2n = 12) híbrido obtido no Brookhaven National Laboratory por intermédio do cruzamento entre a Tradescantia hirsutiflora e a Tradescantia subacaulis. A flor do clone 4430 é azul (dominante) e heterozigota para este locus. Apresenta alta radiossensibilidade, acompanhada por alta sensibilidade a mutágenos químicos também, o que torna este clone especialmente apropriado para estudos de detecção de mutágenos ambientais(26). Dados de radiobiologia demonstraram que os filamentos dos estames do clone 4430 são sensíveis a doses mínimas de 0,25 cGy de raios-X(7,25). Além disso, a Tradescantia foi e tem sido utilizada na monitoração ambiental ao redor de usinas nucleares para a detecção de radionuclídeos que são liberados pelos reatores(27) e no estudo dos efeitos genéticos das radiações ionizantes resultantes do acidente de Chernobyl(28,29) e em testes da mutagenicidade do ar ambiente das regiões da Cracóvia e Polônia após aquele acidente(30). Assim, a Tradescantia pode ser utilizada em estudos controlados sobre os efeitos das radiações ionizantes, bem como servir de sentinela na detecção de vazamentos ou em ambientes de risco. Existem descrições de efeitos sinérgicos da radiação com agentes alquilantes, que não necessariamente se constituem no somatório dos efeitos de cada agente isoladamente(3,31–34). Por esse motivo, a avaliação dos riscos reais deve ser baseada em estudos desses sinergismos entre mutágenos físicos e químicos. Em nosso meio, Saldiva e seu grupo já publicaram estudos sobre os efeitos de poluentes ambientais no clone 4430 da Tradescantia(35). Entretanto, as condições climáticas do Brasil, um país tropical, com calor, umidade e chuvas, se mostraram adversas para o cultivo ideal do clone, com inibição do crescimento e floração das plantas. Além disso, quando cultivada em ambiente externo, sofria ataques constantes de insetos e parasitas, limitando a sua utilização no "mundo real"(36). A Tradescantia pallida var. purpurea, planta da mesma família da Tradescantia (Commelinaceae), que é encontrada com facilidade em canteiros e jardins da cidade de São Paulo, passou a atrair o interesse do grupo. É uma espécie tetraplóide, extremamente resistente a parasitas e insetos, que brota e cresce facilmente, florescendo o ano inteiro. Para checar sua validade como monitor biológico para a genotoxicidade das radiações ionizantes, em relação ao clone 4430, fizeram outro estudo, utilizando o ensaio do micronúcleo(36,37). Neste, a irradiação das plantas demonstrou que a variante popular é igualmente sensível a baixas doses de radiação quanto o clone 4430. Os dados e achados obtidos em experimentos com a Tradescantia são, em geral, consistentes, precisos e confiáveis. Em especial, a resposta a baixos níveis de radiação e mutágenos químicos raramente é obtida com outros ensaios biológicos. Sparrow et al.(38) compararam a radiossensibilidade de diversos organismos, inclusive células de mamíferos, com a da Tradescantia e constataram que os valores de D0 entre 100 e 180 R obtidos com o estudo dos outros organismos se sobrepõem aos valores de D0 obtidos para a Tradescantia em mais de um estudo (149 R, 153 R, 170 R)(2,38). Assim, parece razoável considerar que a Tradescantia apresenta uma radiossensibilidade comparável à das células de mamíferos. As inúmeras semelhanças entre a constituição genética das plantas superiores e dos mamíferos podem levar a crer em efeitos semelhantes de um mutágeno sobre o DNA da planta e do mamífero. Existem, todavia, grandes diferenças organizacionais e fisiológicas entre esses organismos, principalmente na morfogênese e no metabolismo, que podem levar a diferentes reações frente a uma lesão cromossômica. No ser humano, por exemplo, apenas uma fração mínima da lesão sobre o DNA pode ou não levar a mutações, ao passo que na Tradescantia a grande maioria das lesões resulta em mutação. Um determinado aumento na freqüência do dano inicial elevará na mesma proporção a taxa de incidência de mutações, tanto no ser humano quanto na Tradescantia(3,39). Assim, um aumento relativo da freqüência de mutações somáticas na Tradescantia pode indicar um aumento proporcional do risco de mutações para o ser humano.
CONCLUSÕES Os estudos realizados com vegetais apresentam uma série de vantagens que os torna ideais para serem utilizados em países em desenvolvimento(40). • As plantas superiores são eucarióticas — apresentam estrutura cromossômica similar à do ser humano. Passam por mitose, meiose e mutação. É possível o estudo de efeitos sobre a linhagem germinativa comparáveis aos animais. • Podem ser treinados técnicos rapidamente, para a condução de um ensaio com plantas. São relativamente fáceis de cultivar e pouco dispendiosas para se trabalhar. • Algumas apresentam um período germinativo muito curto. • Os estudos podem ser conduzidos sob diversas condições ambientais, pH e temperatura. As plantas superiores podem ser regeneradas de simples células aplóides ou diplóides. • Os ensaios com plantas superiores podem ser utilizados para avaliar a genotoxicidade de simples substâncias químicas até compostos complexos. • Os sistemas de pólen são análogos aos microbiais, em que a graduação dos eventos pode ser feita na grandeza de milhões de células. Podem ser avaliadas aberrações citológicas, mutações genéticas na planta inteira, nas folhas, pólen, embriões, etc. • Os ensaios com plantas superiores podem ser utilizados para a monitoração in situ de poluentes mutagênicos. • A sua utilização é de longa data e são altamente confiáveis. Sua utilidade na pesquisa da mutagênese já está comprovada. • Já são disponíveis resultados de genotoxicidade para uma série de agentes químicos, de forma que podem ser feitas comparações entre diferentes ensaios. • Estudos demonstraram correlação positiva com ensaios citogenéticos em mamíferos. • As plantas superiores podem ser combinadas a ensaios com micróbios para a detecção de metabólitos mutagênicos intermediários como os promutágenos. • Os ensaios de genotoxicidade têm demonstrado alta sensibilidade (raros falso-negativos) na previsão do potencial carcinogênico de diversos agentes. • Centenas de loci genéticos podem ser monitorados. Esse sistema de plantas pode ser empregado tanto em laboratório como em pesquisas de campo, na biomonitoração de inúmeros agentes mutagênicos. É um sistema rápido, pouco dispendioso, versátil e que requer um período de treinamento curto, podendo servir como teste para uma grande variedade de estudos. Com relação às radiações ionizantes, a Tradescantia pallidum, de fácil obtenção e cultivo em nosso meio, poderá ser utilizada como padrão para estudos futuros de mutagenicidade e aberrações cromossômicas, por meio da comparação com os efeitos produzidos por outros tipos de radiação, avaliação do efeito da associação da radiação com drogas ou outros agentes químicos, além da biomonitoração de ambientes de alto risco.
REFERÊNCIAS 1. Read J. Radiation biology of Vicia faba in relation to the general problem. Oxford: Blackwell, 1959; 270–283. [ ] 2. Ichikawa S, Sparrow AH. Radiation-induced loss of reproductive integrity in the stamen hairs of a polyploid series of Tradescantia species. Radiat Bot 1967;7:429–441. [ ] 3. Ichikawa S. Tradescantia stamen-hair system as an excellent botanical tester of mutagenicity: its responses to ionizing radiations and chemical mutagens, and some synergistic effects found. Mutat Res 1992;270:3–22. [ ] 4. Underbrink AG, Schairer LA, Sparrow AH. Tradescantia stamen hairs: a radiobiological test system applicable to chemical mutagenesis. In: Hollaender A, editor. Chemical mutagens: principles and methods for their detection. New York: Plenum Publishing Co, 1973;171–207. [ ] 5. Sparrow AH, Sparrow RC. Spontaneous somatic mutation frequencies for flower in several Tradescantia species and hybrids. Environ Exp Bot 1976; 16:23–43. [ ] 6. Schairer RA, Sautkulis RC. Detection of ambient levels of mutagenic atmospheric pollutants for mutagenicity activity with the higher plant Tradescantia. In: Klekowski EJ Jr, editor. Environmental mutagenesis, carcinogenesis and plant biology. New York: Praeger, 1982;153–194. [ ] 7. Schairer LA, Van't Hof J, Hayes CG, Burton RM, de Serres FJ. Exploratory monitoring of air pollutants for mutagenicity activity with the Tradescantia stamen hair system. Environ Health Perspect 1978; 27:51–60. [ ] 8. Ma TH, Harris MM. In situ monitoring of environmental mutagens. In: Saxena J, editor. Hazard assessment of chemicals: current developments. New York: Academic Press, 1985;83–90. [ ] 9. Sax K. Chromosome aberration induced by X-rays. Genetics 1938;23:494–516. [ ] 10. Sax K. The effect of ionizing radiation on chromosomes. Q Rev Biol 1957;32:15–26. [ ] 11. Sparrow AH. Radiation sensitivity of cells during mitotic and meiotic cycles with emphasis on possible cytochemical changes. Ann N Y Acad Sci 1951;51:1508–1540. [ ] 12. Kirby-Smith JS, Daniels DS. The relative effects of x-rays, gamma rays and beta rays on chromosomal breakage in Tradescantia. Genetics 1953;38:375–388. [ ] 13. Kirby-Smith JS, Graig DL. The induction of chromosome aberrations in Tradescantia by ultraviolet radiation. Genetics 1957;42:176–187. [ ] 14. Ma TH, Snope AJ, Chang TY. Far-red light effect on ultraviolet light induced chromatid aberrations in pollen tubes of Tradescantia. Radiat Bot 1971; 11:39–45. [ ] 15. Ma TH, Isbandi D, Khan SH, Tseng YS. Low level of SO2 enhanced chromatid aberrations in Tradescantia pollen tubes and seasonal variation of aberration rates. Mutat Res 1973;21:93–100. [ ] 16. Ma TH, Khan SH. Pollen mitosis and pollen tube growth inhibition by SO2 in cultured pollen tubes of Tradescantia. Environ Res 1976;12:144–149. [ ] 17. Swanson CP. Effect of ultraviolet treatment on the pollen tube chromosomes of Tradescantia. Genetics 1942;27:491–499. [ ] 18. Khan SH, Ma TH. Hydroxiurea enhanced chromatid aberrations in Tradescantia pollen tubes and seasonal variation of aberration rates. Mutat Res 1974;25:33–37. [ ] 19. Smith HH, Lotfy TA. Comparative effect of certain chemicals on Tradescantia chromosomes are observed at pollen tube mitosis. Am J Bot 1954;41: 589–598. [ ] 20. Ma TH, Kontos GJ, Anderson VA. Stage sensitivity and dose response of meiotic pollen mother cells of Tradescantia to x-rays. Environ Exp Bot 1980; 20:169–174. [ ] 21. Ma TH. Tradescantia micronuclei (Trad-MCN) test for environmental clastogens. In: In vitro toxicity testing of environmental agents. Kolber, Wong, Grant & Hughes, editors. New York: Plenum Publishing Co, 1983. [ ] 22. Ma TH, Cabrera GL, Chen R, et al. Tradescantia micronucleus bioassay. Mutat Res 1994;310:221–230. [ ] 23. Ma TH, Sparrow AH, Schairer LA, Nauman AF. Effect of 1,2-dibromoethane (DBE) on meiotic chromosomes of pollen mother cells of Tradescantia to X-rays. Mutat Res 1978;58:251–258. [ ] 24. Ma TH. Micronuclei induced by X-rays and chemical mutagens in meiotic pollen mother cells of Tradescantia – a promising mutagen system. Mutat Res 1979;64:307–313. [ ] 25. Sparrow AH, Underbrink AG, Rossi HH. Mutations induced in Tradescantia by small doses of X-rays and neutrons: analysis of dose-response curves. Science 1972;176:916–918. [ ] 26. Nauman CH, Sparrow AH, Schairer LA. Comparative effects of ionizing radiation and two gaseous chemical mutagens on somatic mutation induction in one mutable and two non-mutable clones of Tradescantia. Mutat Res 1976;38:53–70. [ ] 27. Ichikawa S. In situ monitoring with Tradescantia around nuclear power plants. Environ Health Perspect 1981;37:145–164. [ ] 28. Ichikawa S, Nakano A, Kenmochi M, et al. Yearly variation of spontaneous somatic mutation frequency in the stamen hairs of Tradescantia clone KU 9 grown outdoors, which showed a significant increase after the Chernobyl accident. Mutat Res 1996;349:249–259. [ ] 29. Shevchenko VA. Some aspects of the genetic consequences of the Chernobyl disaster. In: Hadjian AH, editor. Transactions of the 10th International Conference on Structural Mechanics in Reactor Technology, 1989;vol D:245–249. [ ] 30. Cebulska-Wasilewska A. Tradescantia stamen-hair mutation bioassay on the mutagenicity of radioisotope-contaminated air following the Chernobyl nuclear accident and one year later. Mutat Res 1992;270:23–29. [ ] 31. Cebulska-Wasilewska A, Leenhouts HP, Chadwick KH. Synergism between EMS and X-rays for the induction of somatic mutations in Tradescantia. Int J Radiat Biol Relat Stud Phys Chem Med 1981;40: 163–173. [ ] 32. Knasmuller S, Kim TW, Ma TH. Synergistic effect between tannic acid and X-rays detected by the Tradescantia-micronucleus assay. Mutat Res 1992; 270:31–7. [ ] 33. Nauman CH, Klotz PJ, Schairer LA. Uptake of tritiated 1,2-dibromoethane by Tradescantia floral tissues: relation to induced mutation frequency in stamen hair cells. Environ Exp Bot 1979;19:201–215. [ ] 34. Shima N, Ichikawa S. Synergistic effects of N-ethyl-N-nitrosourea (an alkylating agent with a low Swain-Scott substrate constant) and X-rays in the stamen hairs of Tradescantia clone BNL 4430. Environ Mol Mutagen 1997;29:323–329. [ ] 35. Batalha JRF, Guimarães ET, Lobo DJA, et al. Exploring the clastogenic effects of air pollutants in São Paulo (Brazil) using the Tradescantia micronuclei assay. Mutat Res 1999;426:229–232. [ ] 36. Suyama F, Guimarães ET, Lobo DJA, et al. Pollen mother cells of Tradescantia clone 4430 and Tradescantia pallida var. purpurea are equally sensitive to the clastogenic effects of X-rays. Braz J Med Biol Res 2002;35:127–129. [ ] 37. Carvalho HA, Guimarães ET, Lopes RMG, Lichtenfels AJFC, Lobo DJA, Saldiva PHN. Avaliação de um novo monitor biológico da genotoxicidade das radiações ionizantes. XXIX Congresso Brasileiro de Radiologia, Salvador, novembro de 2000. [ ] 38. Sparrow AH, Underbrink AG, Sparrow RC. Chromosomes and cellular radiosensitivity. I. The relationship of D0 to chromosome volume and complexity in seventy-nine different organisms. Radiat Res 1967;32:915–945. [ ] 39. Ichikawa S. Somatic mutation frequencies in Tradescantia stamen hairs treated with relatively low thermal neutron fluxes. Radiat Res 1997;147:109–114. [ ] 40. Grant WF. The present status of higher plant bioassays for the detection of environmental mutagens. Mutat Res 1994;310:175–185. [ ]
Recebido para publicação em 27/7/2004. Aceito, após revisão, em 3/8/2004.
* Trabalho realizado no Serviço de Radioterapia da Divisão de Oncologia do Instituto de Radiologia (InRad) do Hospital das Clínicas da Faculdade de Medicina da Universidade de São Paulo (HC-FMUSP), São Paulo, SP. |
|

Av. Paulista, 37 - 7° andar - Conj. 71 - CEP 01311-902 - São Paulo - SP - Brazil - Phone: (11) 3372-4544 - Fax: (11) 3372-4554